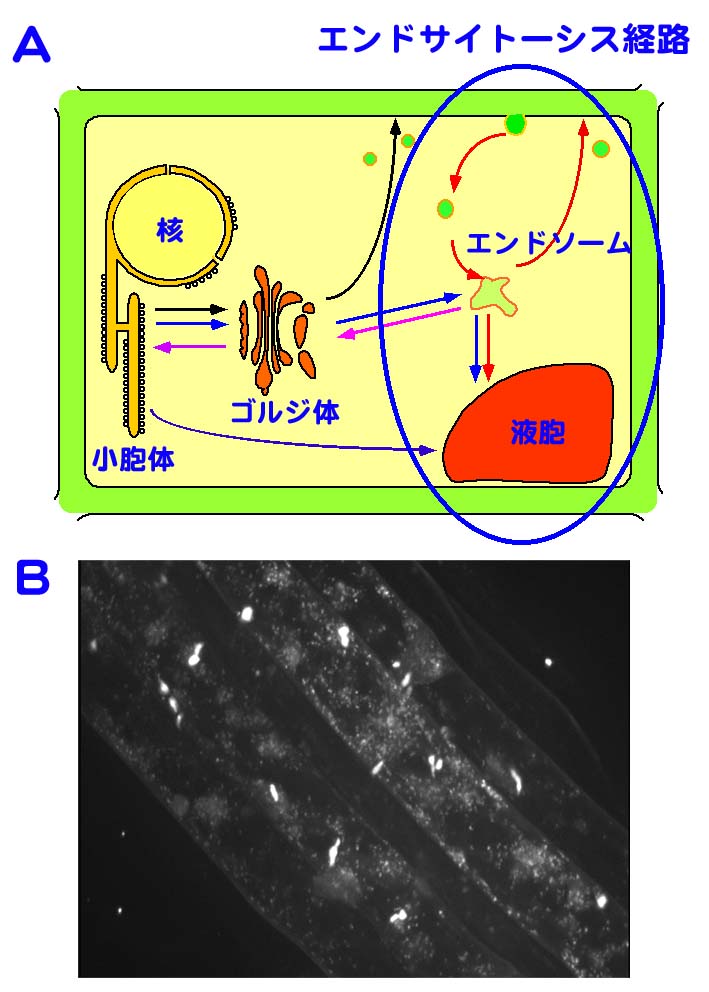
細胞内には,小胞体,ゴルジ体,液胞,エンドソーム等の一重膜により囲まれた細胞小器官(オルガネラ)が数多く存在し,これらの間では,小胞輸送と総称される小さな膜小胞を介した物質輸送機構により,タンパク質や脂質のやりとりが活発に行われている.細胞中に張り巡らされた小胞輸送経路網は,小胞が供与側のオルガネラから出芽,遊離し,目的地となるオルガネラ膜に接着,融合する過程が繰り返されることにより成り立っており,それら各ステップのメカニズムを分子レベルで明らかにしようという研究が,現在様々な生物を用いて精力的に行われている.高等植物においても,小胞体で合成されたタンパク質を液胞へと輸送する経路をはじめとし,小胞体-ゴルジ体間の輸送経路,ゴルジ経由で細胞外へと物質を輸送する分泌経路等の機構が徐々に明らかになりつつある(図 A).我々は,この小胞輸送経路の中でも特に,細胞外のものを細胞内に取り込むエンドサイトーシス経路に注目し,Rab/Ypt GTPaseと呼ばれる低分子量のGTPaseを主な対象として研究を行っている(図 A).Rab/Ypt GTPaseは,輸送小胞が目的地であるオルガネラの膜を認識し,融合する段階で分子スイッチとして機能する分子で,シロイヌナズナのゲノム中には57個のRab/Ypt GTPaseがコードされいる.なかでもAra6は,高等植物に特異的なRab/Ypt GTPaseで,エンドサイトーシスを担うオルガネラであるエンドソームに局在し,その融合を制御している.
我々は,Ara6の機能のさらなる解析を進めるとともに,Ara6をエンドソームのマーカーとして使用することにより,植物細胞内におけるエンドソームのダイナミクスを明らかにすることを目指している.図Bに見られるように,エンドソームは細胞質中の非常に細かいドットとして観察される(やや大きい明るい輝点はプラスチドの自家蛍光).この像は,Ara6と緑色蛍光タンパク質(GFP)のキメラタンパク質を発現する形質転換シロイヌナズナの根の表皮細胞を,高感度・高速共焦点顕微鏡システムを用いて観察したものである.この手法により,エンドソームが細胞中を非常にダイナミックに動き回っていること,その動きがアクチン繊維に依存していること,小胞輸送を阻害することが知られているbrefeldin Aという薬剤が,エンドソームの凝集を引き起こすこと等を明らかにすることができた.今後は,エンドソームと他のオルガネラの相互作用等も含め,植物細胞内におけるオルガネラのダイナミクスをより詳細に解析したいと考えている.