メリステム形成・維持に必須な細胞接着関連遺伝子の同定と機能解明
佐藤 忍(筑波大学生物科学系)
研究協力者:岩井宏暁(学振特別研究員)
茎頂や根端などのメリステムにおいては、細胞は高い分裂活性を維持しつつ周囲の細胞群と常に接し、緊密な情報交換を行っている。そのためには、強い細胞同士の接着が必須である。しかし現在のところ、細胞接着の主役であるペクチンの生合成機構に関する知見は極めて乏しい。本研究では、半数体タバコを用いた新たな変異体作出系により細胞接着変異体を作出し、その遺伝子を解析することにより、高等植物の細胞接着に関わる遺伝子のメリステム形成と維持における役割を明らかにする事を目的としている。
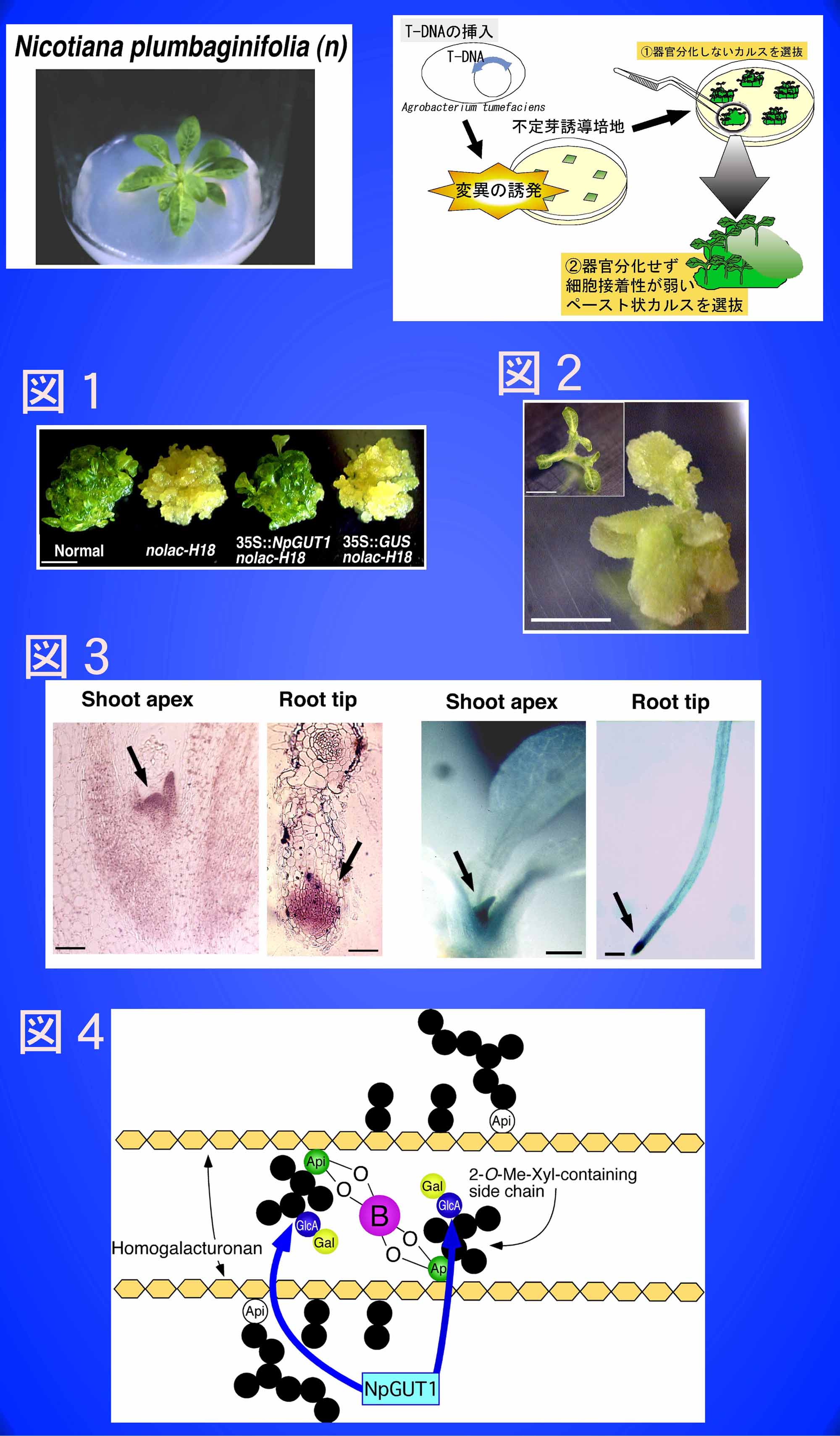
半数体Nicotiana plumbaginifoliaの葉切片にアグロバクテリウムを介してT-DNAを導入し、不定芽形成能力の喪失と共に細胞接着性の低下した変異体を作出した。そのうちnolac-H18株において、変異の原因遺伝子としてペクチン合成としては初めてのNpGUT1 (glucuronyltransferase 1) が同定された。
図1
Normal、nolac-H18変異体、NpGUT1相補体の形態
nolac-H18の相補形質(不定芽を形成せず、細胞接着性が弱い)は、NpGUT1の過剰発現により相補された(35S::NpGUT1 nolac-H18)。右端はGUSを過剰発現させた対照(スケールは5 mm)。
図2
NpGUT1アンチセンス植物体の形態
NpGUT1の発現を抑制すると、組織間の接着が非常に弱い形質転換体が形成された。
細胞壁の強度が弱くなっただけでなく、NpGUT1の発現が強いメリステム内での細胞接着が弱くなった結果、原形質連絡が絶ちきられ、緊密な情報交換が不全になり正常な分化ができなくなったと考えられる。
図3
NpGUT1をプローブとしたin situハイブリット形成(左側、スケールは50 μm)とpNpGUT1::GUS形質転換体(右側、スケールは5 mm)。
NpGUT1は、タバコの茎頂および根端のメリステムで強く発現している。
図4
ペクチンRG-II領域は、一分子のホウ素とエステル結合することでペクチン分子間を架橋している。
NpGUT1は、ホウ素-ペクチン架橋に必須である側鎖のグルクロン酸(GlcA)を転移する酵素である。nolac-H18変異体では、GlcAが転移されなかったことでガラクトース(Gal)も同様に転移されていなかった。その結果、GlcA-Galの2糖が欠落し、ホウ素ペクチン架橋形成能力を失っていた。