サイトカイニンは植物の組織形成および細胞の増殖・分化において中心的な役割を果たす植物ホルモンである。細胞レベルにおいては、オーキシンと共役的に働き増殖を促進する。一方、組織レベルにおいては、維管束の分化、側芽形成、葉の成長拡大を促進するとともに、老化を遅らせる。また、茎頂分裂組織の発達・維持に関しても、サイトカイニンは細胞の増殖・分化の制御を通じて重要な役割を担っていると考えられる。このように、サイトカイニンのもつ作用は多様であるが、それらの作用がもたらされるための分子機構については、ほとんど解明されていない。
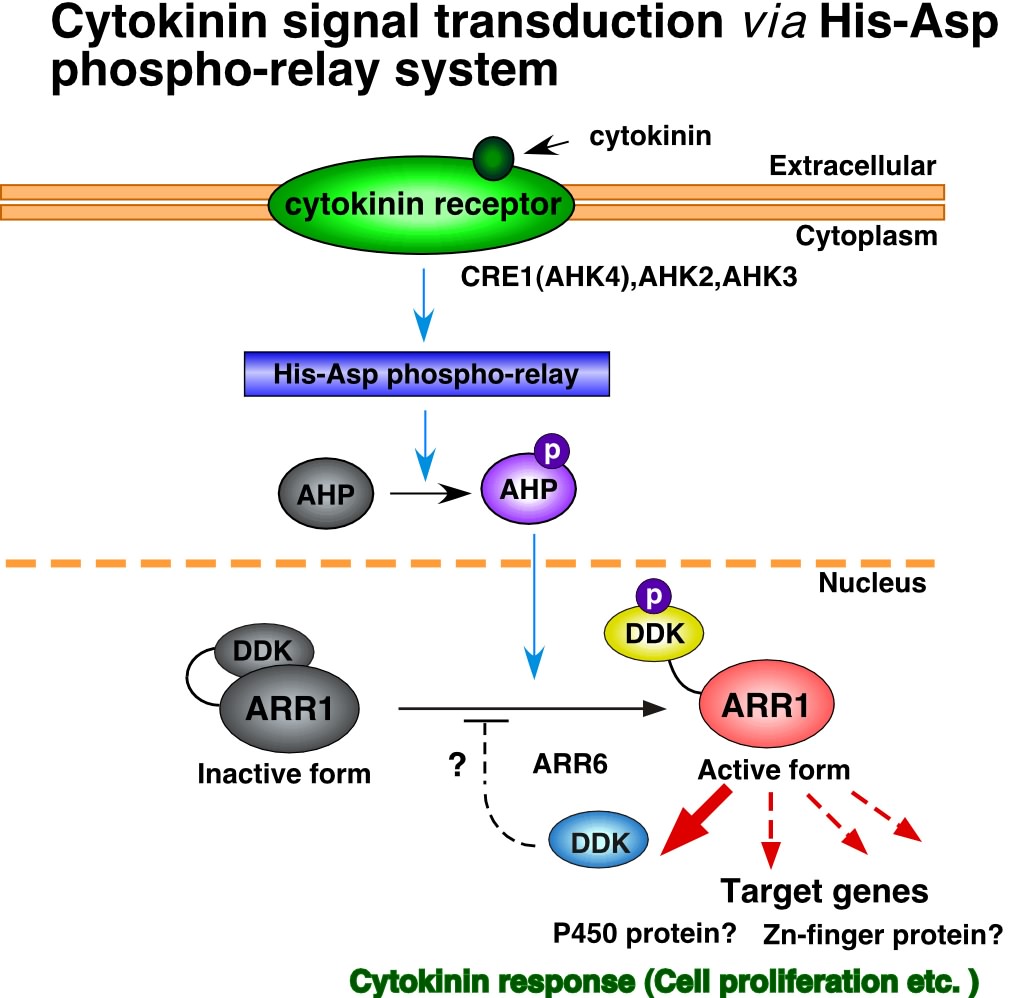
サイトカイニンの研究分野は、この数年の間に飛躍的な進歩を遂げた。サイトカイニン受容体がHis-Aspリン酸リレーに関与するHisキナーゼであることの発見や、植物体内でサイトカイニン合成の鍵となる酵素が同定されたことなどがあげられる。さらに、我々のグループはシロイヌナズナのHis-Aspリン酸リレーに関与する転写因子型レスポンスレギュレーターARR1およびARR2が、サイトカイニン受容体からのリン酸リレーの下流に位置し、サイトカイニンの初期応答遺伝子であるARR6の転写を直接活性化することを明らかにした。これによってサイトカイニンは植物ホルモンとして初めて、受容から初期応答遺伝子に至るシグナル伝達経路の概容が明らかにされた。しかし、標的遺伝子として同定されたARR6はレシーバードメインに続く伸長領域をもたないレスポンスレギュレーターであり、その分子機能についても今のところ明らかにされていない。サイトカイニン存在下でARR1遺伝子を過剰発現させた植物体を生育させると、茎頂や子葉、胚軸において異所的なシュートやカルス形成が誘導されることから、ARR1の標的遺伝子はARR6以外にも多数存在し、細胞増殖や細胞分裂に関わる遺伝子が標的となっている可能性が高い。そこで、ARR1の転写因子活性を誘導的に制御できる系を用いてcDNAマイクロアレイ法により、ARR1の標的遺伝子の探索を行った。転写レベルの上昇が著しかった遺伝子の中にはAタイプのレスポンスレギュレーター遺伝子ARR5, ARR6, ARR7, ARR16, ARR17が含まれていた。この他にも、P450タンパク質およびZinc fingerタンパク質が有意に多く見いだされた。これらの候補がすべてARR1の標的遺伝子とは考えがたいが、Aタイプのレスポンスレギュレーター遺伝子が多数含まれていることから、ある程度の濃縮はかかっていると期待される。今後、これらが標的遺伝子であるかどうかを調べることによって、ARR1が関与するサイトカイニンシグナル伝達の概要が明らかにされると考えられる。また、サイトカイニン受容体であるCRE1(AHK4)の変異体(wol、cre1-1)では、根の維管束の細胞列の数が減少し、師管、道管の分化が正常におこらない。ARR1がサイトカイニン受容体CRE1の下流で機能しているかどうかを調べるために、ARR1の転写因子活性を構成的にあるいは誘導的に制御できる系をwol、cre1-1変異体に導入した形質転換植物体を作成し、これらの変異体の表現型をARR1がsuppressできるかどうか調べた。wol変異体にARR1の転写因子活性を構成的に制御できる系を導入した植物体では少なくとも根の伸長阻害の回復が観察された。このことから、ARR1はCRE1の下流で機能している可能性が高いことが示唆された。