花序のメリステム形成をエピジェネティックに抑制するTFL2遺伝子の解析
岡山県生物科学総合研究所 後藤弘爾
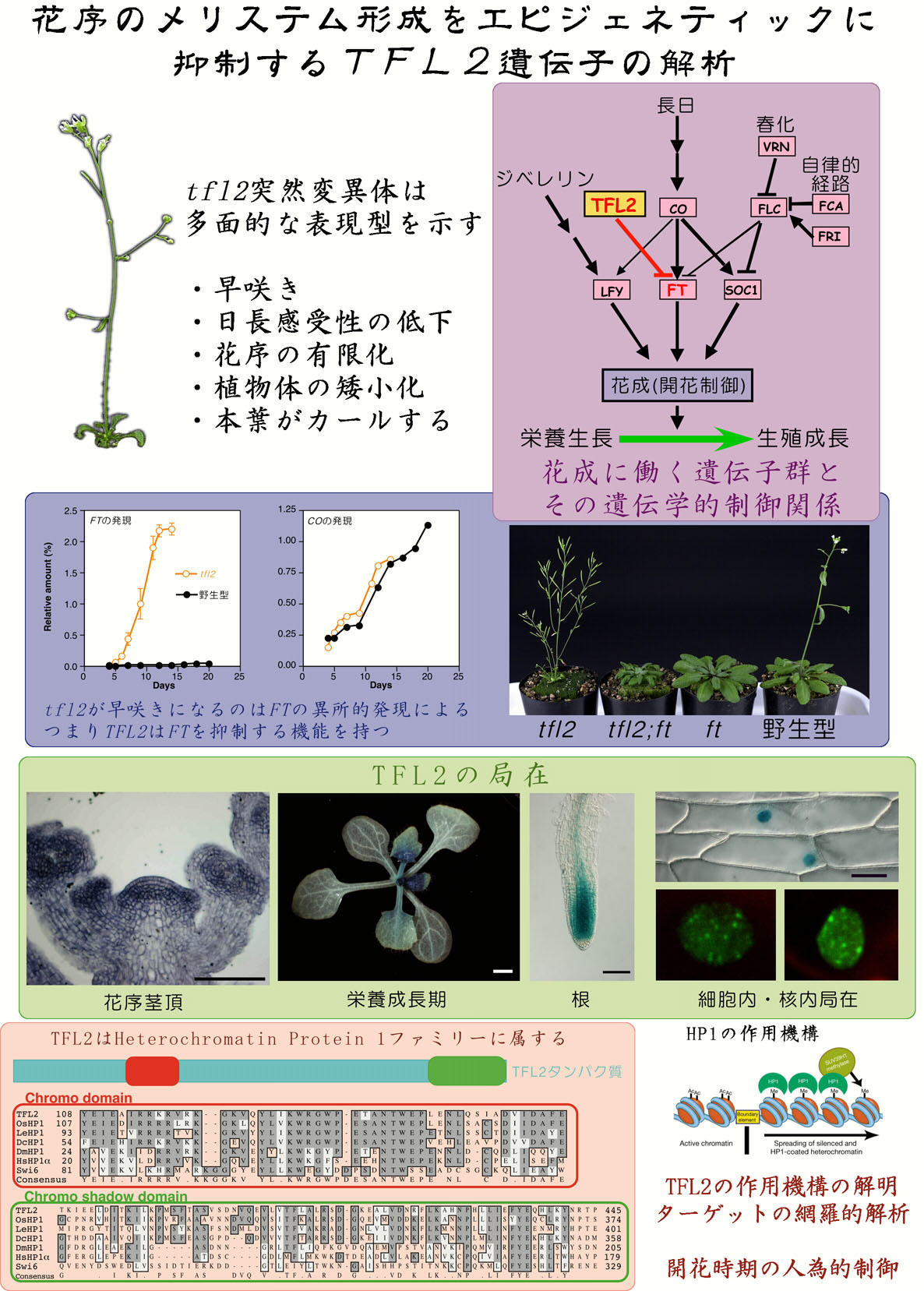
植物が花芽を分化する現象、すなわち栄養生長から生殖性長への転換を花成と呼ぶ。モデル植物、アラビドプシス(シロイヌナズナ)を用いた分子遺伝学的研究から、花成に関わる遺伝学的な経路が明らかになってきた。これらの研究は主に遅咲きになる突然変異体を用いた、花成を促進する遺伝子の研究が中心であった。そこで私たちは早咲きになる突然変異体であるtfl2に着目し、花成を抑制する遺伝子を研究することで、これまでに明らかになった花成の遺伝子とは異なったカテゴリーの制御機構を明らかにすることが出来ると考えた。
遺伝子発現の変化を野生型とtfl2とで比較したところ、花成関連の遺伝子ではFT遺伝子の発現量だけがtfl2で著しく上昇していた。FT遺伝子は長日刺激を受けて花成を促進することが知られているので、tfl2が日長条件に関係なく早咲きになることとも合致する。特にFTを直接転写活性化するCO遺伝子の発現が野生型とtfl2とで変化しないことから、TFL2はFT特異的な抑制因子であるといえる。これは遺伝学的にも、二重突然変異体の解析によって確かめられた。
さて、TFL2であるが、遺伝子クローニングによって動物のヘテロクロマチンタンパク質(HP1)のホモログをコードしていることが明らかとなった。HP1は染色体上で遺伝子が不活性な領域、ヘテロクロマチン領域に多量存在し、その形成と維持に関与していると考えられている。TFL2もその構造や植物細胞の核内に点状に局在すること、分裂酵母のHP1(Swi6)突然変異を相補することなどから、HP1と同様な機能を持つと推測される。またtfl2の多面的な表現型も考え合わせると、TFL2はエピジェネティックな制御に関与していると予想される。しかしながら、FTはユークロマチン遺伝子であることや、tfl2ではヘテロクロマチン領域の遺伝子発現が活性化されていないなど、HP1で分かったことをそのままTFL2に当てはめることが出来ないということも明らかとなった。
本研究でこれまでに得られた以上のような結果を踏まえて、今後私たちはTFL2がどの様なメカニズムで植物細胞内において遺伝子抑制を行っているのかを明らかにしていく計画である。
研究室ホームページ:http://ns.bio-ribs.com/~goto-lab/
エピジェネティック: もともと「後生的」を意味する言葉「エピジェネティック」は、現在では遺伝子の塩基配列に違いがないのに、その遺伝子活性の違いによって引き起こされる生命現象を指す場合が多い。DNAのメチル化による発現制御がよく知られた例である。このようなエピジェネティックな制御は複数の遺伝子に対して起きることが多く、その表現型は多面的であることが多い。