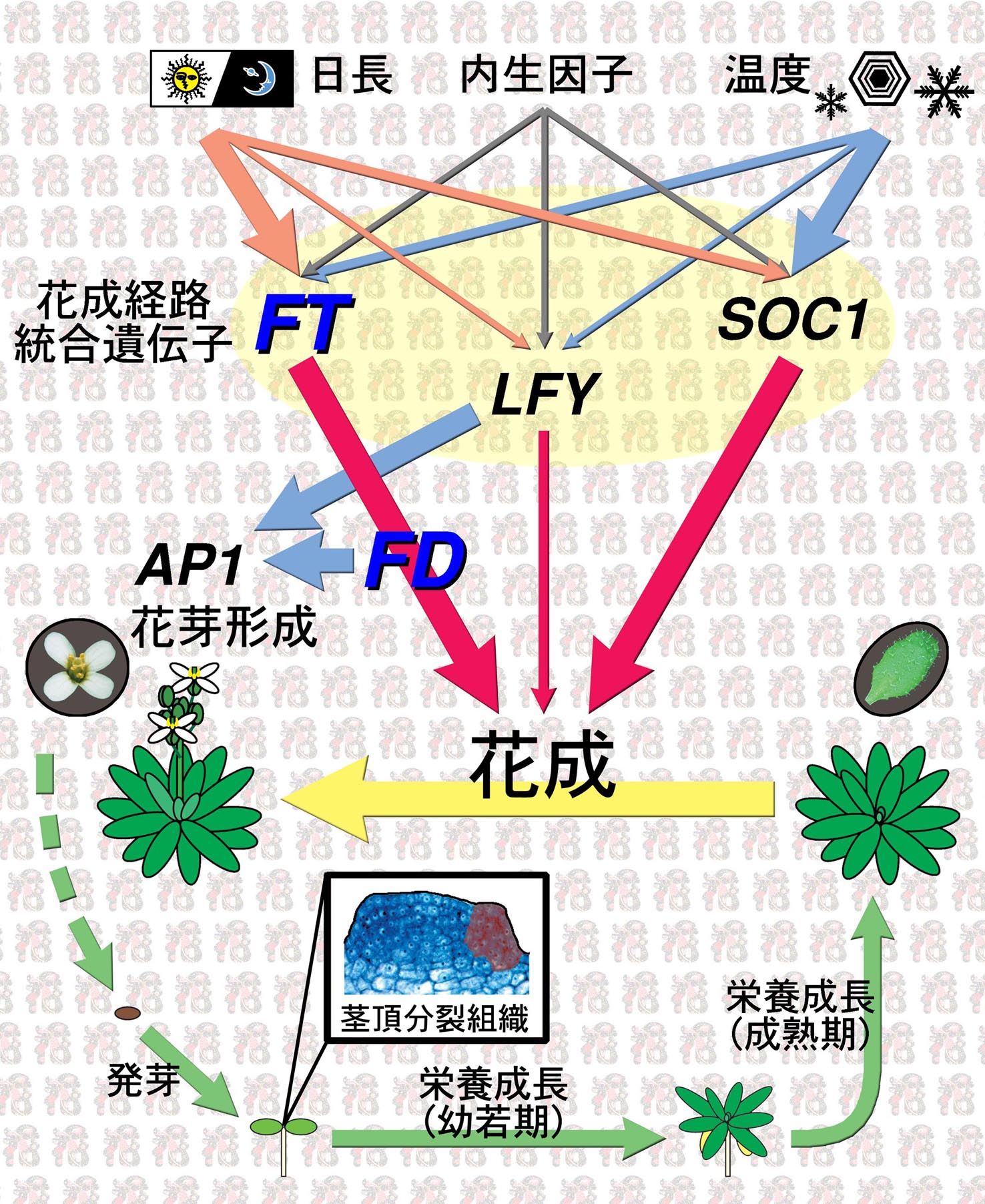
シロイヌナズナ(一年生草本植物)の生活環と花成制御
発芽後、幼植物は2枚の子葉の間にある茎頂分裂組織(枠内。赤く塗った部分が器官原基が形成される領域)でおこなわれる器官形成により地上部の全構造を形成する(後胚発生)。茎頂分裂組織は、まず栄養成長相と呼ばれる成長相に入る。この間に形成される器官原基はもっぱら葉原基であり、葉に分化する(右の楕円内)。植物は、しばらく栄養成長を続けた後、花成 (floral transition)とよばれる成長相の転換を経て、生殖成長相に移行する。生殖成長相では、茎頂分裂組織は葉原基に代わって花芽原基を形成するようになる。花芽原基は花に分化する(左の円内)。
花成は、植物個体の"齢"などの内的要因と外的な環境要因による複雑な制御を受ける。環境要因のうち日長(光周期)と温度が特に重要なものとして知られている。日長については、シロイヌナズナでは長日条件が花成に促進的である。短日条件が花成を促進するイネやアサガオとは対照的である温度については、シロイヌナズナを含む多くの植物で、冬に経験する長期間の低温被曝の効果が重要であることが知られており、春化 (vernalization)とよばれる。
われわれの研究グループを含む多くのグループの研究により、これらの内的・外的要因からの信号は異なる制御経路を経て、最終的には、"花成経路統合遺伝子"(floral pathway integrator)と呼ばれる少数の遺伝子の転写制御の段階で統合されることが明らかになってきた。花成経路統合遺伝子としては、われわれの研究グループがクローン化し解析を進めてきたFT遺伝子の他に、SOC1遺伝子、LFY遺伝子が知られている。FT遺伝子産物は、哺乳動物のフォスファチジルエタノールアミン結合蛋白質(Raf-1キナーゼ阻害蛋白質)と高い相同性を持つが、その生化学的な機能は不明である。われわれの研究から、FT遺伝子産物による花成の実行にはbZIP型の転写因子をコードする花成遺伝子FDの機能が必要であることが明らかになった。一方、FD蛋白質の機能発現にはFT遺伝子機能が必要であることも明らかになった。FT蛋白質とFD蛋白質の間には相互作用がみられることから、蛋白質間相互作用を介してFT蛋白質がFD蛋白質の活性を調節していると考えられる。この活性調節の実態を明らかにすることは重要な課題のひとつである。転写因子FDの標的遺伝子は未同定であるが、有力な候補のひとつに、花芽形成の初発過程に関わるAP1遺伝子がある。FDは他の転写因子LFYとともに花芽分裂組織におけるAP1遺伝子の転写を活性化すると考えられる。FDの標的遺伝子はまだ多数存在すると考えられ、それらの同定は花成の実行過程を明らかにする上で重要な課題である。